
新概念を学ぶ13 原猿の本能不全によって性闘争が抑止され、扁桃体の仲間認識(追従本能)が解除された

前回「新概念を学ぶ12~哺乳類と原猿の集団化の違い」で扱ったのは、『実現論』「前史ニ.サル時代の同類闘争と共認機能」のうちの以下の部分である。
原猿たちは恒常的に飢えの苦痛に苛まれ、いつ襲ってくるか分からない敵=首雄の攻撃に怯えながら暮らしていたが、それらの極度な不全感が生命の根源を成す適応欠乏を強く刺激し、生起させた。加えて、恒常的に強力な危機逃避回路(未解明だが、おそらくアドレナリンetc.の情報伝達物質)が作動する事によって(これも未解明だが親和系のオキシトシンetc.による性封鎖力ともあいまって)性闘争が抑止され、それによって、モグラ以来性闘争物質によって封鎖されてきた追従本能が解除された。
かくして、不全感の塊であった境界空域の弱オスたちは、適応欠乏に導かれて強く追従本能に収束する。しかし、互いに追従し合っても、誰も(縄張りの確保あるいは不全感の解消の)突破口を示すことは出来ない。そこで、わずかに可能性が開かれた(=不全感を和らげることのできる)親和本能を更に強化し、追従回路(アドレナリンetc.)に親和回路(オキシトシンetc.)が相乗収束した依存本能に収束してゆく。つまり、「縄張りを持たない敗者たちが互いに身を寄せ合う」。
このように、原猿は、本能不全⇒適応欠乏と危機逃避回路作動⇒性闘争抑止⇒追従本能+親和回路が相乗収束した依存本能に収束した。
この中で、危機逃避⇒追従回路を司るのが、脳内の扁桃体という機能(部位)である。
今回は、この扁桃体が果たす機能を明らかにする。
いつも応援ありがとうございます。
『るいネット』「【仮説】扁桃体の危機察知・仲間認識回路に、ミラーニューロンによる同一視回路が塗り重ねられ、共感回路(共認原回路)が形成されたのでは?」
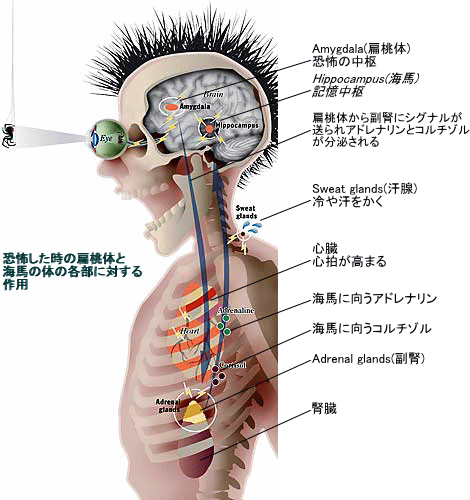
扁桃体の歴史は古く、魚類の段階で扁桃体に相当する領域が形成されている。その機能は多々あるが、その中心機能は「危機察知→逃避行動の発令」と「仲間認識」にある。
危機に面した時、扁桃体(扁桃核)は過去の記憶との照合を行い、自らに危機を与えるものと判断が行われると、神経伝達物質の分泌によって、脳と身体を危機に備えるように指令する。
一方で、扁桃体は仲間認識や社会性の形成にも関わっており、魚類段階でも扁桃体に相当する領域が仲間を認識し、群れを形成する動因となっていることが解っている。
これらのことから、「危機察知」と「仲間認識」は一体であり、脊椎動物の進化過程で、「仲間認識」によってより高度な「危機察知→逃避」回路が形成されたと言うことが解る。
なお、この「危機察知」と「仲間認識」を行うのは、主要に右脳の扁桃体であることが解っている。
右脳・左脳の機能分化の中で、特に右脳は危機察知・危機対処と仲間認識の機能を分担して進化してきたことが解っており、扁桃体をその中心核として、右脳が危機察知・仲間認識脳として進化してきたと言える。
また、扁桃体は、危機察知・仲間認識を行う上で、快・不快、有益・有害などの情動判断を行っており、情動反応に基づいて多様な神経伝達物質を分泌、脳の興奮と抑制を制御する。

扁桃体の機能は脊椎動物(魚類)から登場したのは何故か?
それ以前の軟体動物や節足動物に比べて飛躍的に運動量(速度)を上昇させたのが脊椎動物(魚類)である。
この進化の主圧力は、外敵圧力(種間闘争圧力)である。
脊椎動物は外敵圧力(種間闘争圧力)に適応するために、脳~脊椎という中枢指令機能を集約することによって、外敵からの危機察知⇒逃避行動or餌の察知⇒捕食行動を大幅にスピードアップさせた。
中枢指令機能の中の扁桃体も、この外敵圧力(種間闘争圧力)に適応するために形成された機能であり、外敵か餌か等のプラスマイナス判断を即時に行う機能(部位)として登場したと考えられる。
つまり、扁桃体は同類以外の対象は外敵or餌と認識し、危機逃避or捕食する。
また、生物は危機逃避するために群れ集まるが、脊椎動物は扁桃体によって、同類に対する仲間認識を強化するとともに、(単に群れ集まるだけではなく)集団としての危機逃避行動を飛躍的に高度化orスピードアップさせた。つまり、扁桃体の危機察知・仲間認識機能が、追従本能を強化・高度化したと考えられる。
ところが、性情動(性闘争)物質が働くと、扁桃体の仲間認識が封鎖され、同類は性闘争の対象と認識され、性闘争のスイッチが入る。これが性情動(性闘争)物質によって追従本能が封鎖される仕組みだと考えられる。
性闘争本能は、脊椎動物の前から殆どの動物に存在しているが、哺乳類は、淘汰適応の必要から性闘争の本能を極端に強化し、その性情動物質によって追従本能を封鎖することによって、個間闘争を激化させ淘汰を促進するという淘汰促進態である。
その結果、原モグラは追従本能が封鎖され、バラバラに隠れ棲み、性闘争を行うようになったのである。
ところが、原猿が逃避した樹上には外敵が殆どいない。その上、樹上には栄養価の高い果実や木の実が沢山ある。従って、原猿にとっては、自然圧力や外敵圧力(種間闘争圧力)が主圧力ではなく、同類闘争(性闘争)が主圧力に変わっている。危機察知の対象は専ら同類になると云ってもよい。
さらに、原猿の弱雄では、本能不全⇒適応欠乏と危機逃避回路作動によって(親和物質=オキシトシンによる性封鎖力ともあいまって)性情動物質が抑止される。モグラ以来性闘争物質によって封鎖されてきた扁桃体の仲間認識が復活し追従本能が解除された。かくして、不全感の塊であった境界空域の弱オスたちは、適応欠乏に導かれて強く追従本能に収束する。
しかし、互いに追従し合っても、誰も(縄張りの確保あるいは不全感の解消の)突破口を示すことは出来ない。そこで、わずかに可能性が開かれた(=不全感を和らげることのできる)親和本能を更に強化し、追従回路に親和回路が相乗収束した依存本能に収束してゆく。
これは、一般動物から見て異常な事態である。
∵一般哺乳類では成体の雄は親和行為をしないが、原猿の弱雄たちは不全感を和らげるために親和物質(オキシトシン)が分泌され、「縄張りを持たない敗者たちが互いに身を寄せ合った」。また、一般の雌雄分化した生物では雌と子供が依存存在、雄が庇護存在であるが、原猿では弱雄が全て(庇護存在でなくなり)依存存在となってしまう。
この一般哺乳類には見られない弱雄同士の親和行為と弱雄の依存存在化が、親和本能の更なる強化と依存本能への収束の中身である。
この段階でも、弱雄たちは首雄を恐れているが、それは首雄に縄張り闘争で負けた体験記憶に基づいて、弱雄の扁桃体が首雄を「怖い」と判断しているのであろう。
まとめると、
【1】扁桃体の危機逃避回路・仲間認識回路が追従本能の中核を成す。
【2】扁桃体の仲間認識を性情動物質が抑止することで性闘争が起きる。
【3】ところが本能不全に陥った原猿は、適応欠乏と危機逃避回路が作動して性情動物質が抑止された結果、扁桃体の仲間認識が復活し、追従本能が復活した。
【3】ところが、それだけでは本能不全を突破できないので、親和本能をさらに強化した。
その中身は、成体雄にも親和物質(オキシトシン)が分泌され、元々は庇護存在(闘争存在)である弱雄同士が親和し合うという、一般生物からすれば異常事態である。
これが「親和本能を更に強化し、追従回路に親和回路が相乗収束した依存本能に収束してゆく。つまり、縄張りを持たない敗者たちが互いに身を寄せ合う」ということの中身である。
ところが、それでも本能不全は克服されたわけではない。
最終的に、原猿の弱雄たちは、この不全感をどのようにして突破したのだろうか?
次回、明らかにしてゆきます。

トラックバック
このエントリーのトラックバックURL:
http://blog.nihon-syakai.net/blog/2013/06/2553.html/trackback